高 娜,卢 霞,闫允君,孔 杰,孟宪红,李旭鹏,隋 娟,陈宝龙,曹宝祥,高 焕,栾 生
( 1.江苏海洋大学 海洋科学与水产学院,江苏省海洋生物技术重点试验室,江苏 连云港 222005; 2.中国水产科学研究院 黄海水产研究所,农业农村部海洋渔业可持续发展重点实验室,山东 青岛 266071; 3.青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,山东 青岛 266071; 4.中国科学院 烟台海岸带研究所,沿海生物资源高效利用研究开发中心,山东 烟台 264003 )
动物肌肉的发生过程主要处于胚胎发育时期内,是在一系列基因复杂而精确的调控下进行的[1-2]。近年来,DNA重组、基因敲除等多种技术的广泛应用,使得胚胎肌肉生成的分子调控机制研究取得了极大的进展。转化生长因子-β(TGF-β)是一类庞大的调控动物肌肉生长发育的超家族,该家族中的多个基因已被证实与肌肉发育有直接或间接的关系。骨形态发生蛋白(BMP)是从牛骨中分离的TGF-β超家族中的一员[3],可以启动从软骨形成到头骨形成的过程[4]。截至目前,骨形态发生蛋白已被证明是一种多功能分泌信号蛋白,对各种细胞类型包括单核细胞[5-7]、上皮细胞[8]、间充质细胞[9]、神经细胞[10]均有广泛的生物学活性。骨形态发生蛋白调节这些细胞的生长分化,并在各种组织和器官的形态形成中发挥关键作用[11]。对脊椎动物骨形态发生蛋白家族的研究较多,如:褚建新等[12]发现,给正常小鼠大腿肌肉内植入rh-BMP-2基因后,可以激活自身肌肉间质细胞间接拯救骨髓衰竭;有研究证明,BMP-2基因间接参与了爪蟾(Xenopus)肌肉发生的起始特化过程[13]。近年来,一些无脊椎动物如棘皮动物[14-15]、节肢动物[16-17]、软体动物[18-20]等的骨形态发生蛋白家族成员也逐渐被克隆成功,发现其功能与脊椎动物的功能相似。
中国明对虾(Fenneropenaeuschinensis)是黄、渤海区域的特有种类,肉质鲜嫩,具有较高的经济价值,历史最高产量达到2.0×105t;但由于过度捕捞和环境恶化,其野生资源急剧减少,国内养殖产量远远不能满足市场的需求[21-23]。生长速度是决定养殖产量最为重要的经济性状之一。骨形态发生蛋白是动物肌肉生长发育的重要调节因子,在脊椎动物及某些无脊椎动物的生长细胞中起关键作用。然而,关于骨形态发生蛋白调控中国明对虾肌肉发生和生长的研究尚未见相关报道。笔者克隆了中国明对虾骨形态发生蛋白家族中的BMP-7基因成员,通过实时荧光定量PCR (qRT-PCR)技术分析了FcBMP-7基因在中国明对虾幼体不同发育时期和生长速度差异显著的幼虾不同组织中的表达规律,为进一步解析中国明对虾肌肉发生发育的分子机制提供理论依据。
1.1 试验材料
试验所用中国明对虾样本均来自农业农村部(青岛)海水养殖遗传育种中心第13代选育群体。中国明对虾幼体不同发育阶段取样:分别从胚胎发育阶段的原肠期和4个幼体发育阶段无节幼体期、溞状幼体期、糠虾幼体期和仔虾期采集10尾家系样本,每个阶段每个家系取50~60尾混合为1个样本,其中大部分内容物为肌肉。取幼虾期同一家系的2种规格[(3.95±0.08) g和 (1.55±0.12) g]各10尾虾的不同组织部位(心脏、鳃、肝胰腺、胃、肠道、肌肉、步足和游泳足),放入冻存管立即加入液氮后-80 ℃冰箱保存备用。
1.2 试验方法
1.2.1 总RNA的提取与cDNA的合成
利用Trizol法提取以上样品的总RNA,通过1%琼脂糖凝胶电泳检测其完整性,用分光光度计检测RNA浓度。采用HiScript ⅢRT SuperMix for qPCR (+gDNA wiper)逆转录试剂盒(Vazyme,南京)合成第1链cDNA,-20 ℃保存。
1.2.2 FcBMP-7基因的克隆
在中国明对虾的转录组测序数据筛选到BMP-7基因部分cDNA片段。根据已知序列设计5′RACE和3′RACE特异性引物,设计原则参照5′RACE System for Rapid Amplification of cDNA Ends version 2.0 (Invitrogen,美国)和SMARTer®RACE 3′ Kit (Clontech,美国)试剂盒的说明书(表1)。
1.2.3 FcBMP-7蛋白的生物信息学分析
利用BioEdit软件将3′RACE和5′RACE扩增片段与已知序列进行拼接,获得FcBMP-7基因的cDNA全长序列。通过DNAMAN 8.0翻译氨基酸序列;利用BLASTP (http://blast.ncbi.nlm.nih.gov/Blast.cgi)进行预测蛋白序列的同源性比对;通过Protparam (http://www.expasy.org/tools/protparam.html)预测蛋白的等电点和分子质量;采用NetNGlyc 1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc/)查找糖基化位点;通过NetPhos 2.0 Server (http://www.cbs.dtu.dk/services/NetPhos/)查找磷酸化位点;利用ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/)进行氨基酸序列的比对;基于MEGA 7.0的邻接法构建系统进化树(自展值为1000)。
1.2.4 FcBMP-7基因的表达模式分析
以18S rRNA为内参基因,通过实时荧光定量PCR检测FcBMP-7基因在不同发育时期、不同生长速率的2组虾的不同组织中的转录水平,反应体系和反应条件参照ChamQ Universal SYBR qPCR Master Mix试剂盒(Vazyme,南京)说明书进行。利用2-△△Ct方法[24]计算FcBMP-7基因的相对表达量。试验所用引物均由生工生物工程(上海)股份有限公司合成(表1)。
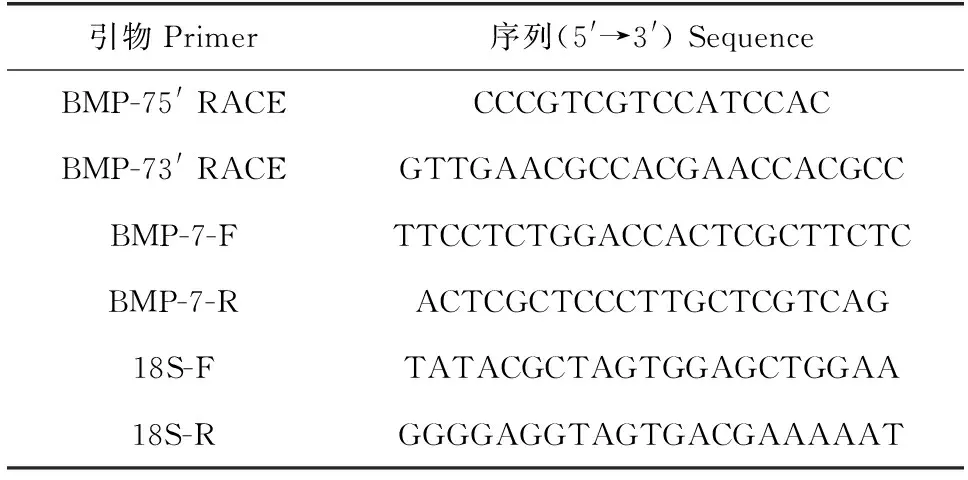
表1 试验所用引物序列Tab.1 The primer sequences used in this study
利用Excel 2016软件分析中国明对虾幼体和幼虾不同组织FcBMP-7基因的表达量。利用SPSS 25.0中的单因素方差分析,对FcBMP-7基因在幼体不同时期间以及幼虾不同规格间的转录表达差异进行显著性检验。
2.1 FcBMP-7基因的克隆及生物信息学分析
由于中国明对虾转录组筛选得到的BMP-7基因cDNA序列不完整,因此分别进行5′RACE和3′RACE扩增5′和3′端的序列。其中5′RACE获得541 bp序列 (图1a),3′RACE得到409 bp序列 (图1b)。与原序列拼接后得到中国明对虾BMP-7基因2003 bp的全长cDNA序列,其中开放阅读框为1242 bp,编码413个氨基酸,5′非编码区为466 bp,3′非编码区为295 bp(图2),将其命名为FcBMP-7基因(GenBank登录号: MW048584)。

图1 RACE扩增片段的电泳结果Fig.1 Eelectrophoresis of RACE amplificationa.5′ RACE电泳结果;b.3′ RACE电泳结果.a.electrphoresis of 5′ RACE amplification; b.electrphoresis of 3′ RACE amplification.

图2 FcBMP-7基因的全长cDNA及推测编码的氨基酸序列Fig.2 Full length cDNA and the putative amino acid sequence of FcBMP-7 gene黑色加粗部分为起始密码子和终止密码子;灰色阴影为磷酸化位点;实线方框为N-糖基化位点;下划线为跨膜螺旋结构;斜体加粗的部分为TGF-β多功能肽.The initiator and the terminator are marked by black bold; the phosphorylation sites are denoted by gray shadow; the N-glycosylation sites are framed with solid lines; a transmembrane helix region is described as underlined part; the multifunctional peptide of TGF-β is expresse by italic bold.
该蛋白的等电点为6.20,分子质量为45.6 ku。蛋白功能结构域预测表明,在该基因编码蛋白序列中,α螺旋占22.28%、延伸链占21.07%、β转角占6.05%、无规则卷曲占50.60%(图3)。

图3 FcBMP-7蛋白的二级结构预测Fig.3 The prediction of the secondary structure for FcBMP-7 proteinConf.预测可信度;Cart.三态分配;Pred.三态预测;AA.目标序列.Conf.confidence of prediction; Cart.3-state assignment cartoon; Pred.3-state prediction; AA.target sequence.
2.2 FcBMP-7蛋白的同源性比对及系统进化树分析
FcBMP-7蛋白的氨基酸序列与其他物种进行同源性比对分析结果见图4。FcBMP-7蛋白与节肢动物各物种的BMP-7蛋白具有很高的相似性,在相对保守区域有7个半胱氨酸残基,其位置分别是307、336、340、377、378、410、412位氨基酸。此外,该蛋白还含有1个跨膜螺旋结构域(13~35位氨基酸)和1个信号肽,信号肽的剪切位点位于第31和32位氨基酸之间。FcBMP-7蛋白与凡纳滨对虾(Litopenaeusvannamei)BMP-7蛋白的同源性最高,达到98%;其次是脊尾白虾(Palaemoncarinicauda)和中华绒螯蟹(Eriocheirsinensis),同源性分别是75%和72%;与软体动物如文蛤(Meretrixmeretrix)及三角帆蚌(Hyriopsiscumingii)等的相似性较低,同源性为30%~34%。

图4 FcBMP-7蛋白序列与其他已知物种BMP-7蛋白的同源比对结果Fig.4 Results of homologous comparison between FCBMP-7 protein sequence and BMP-7 protein of other known speciesBMP-7氨基酸序列比对物种的GenBank登录号:凡纳滨对虾BMP-7, XM_027355389.1;脊尾白虾BMP-7, AYC12382.1;中华绒螯蟹BMP-7, AWV66904.1;拟穴青蟹BMP-7, AOZ56940.1;智人BMP-7, NM_001719.3;原鸡BMP-7, XM_417496.6;莫桑比克罗非鱼BMP-7, FN565169.2;非洲爪蟾BMP-7, NP_001079934.1.GenBank accessions numbers of BMP-7 amino acid sequence alignment:L. vannamei BMP-7, XM_027355389.1; P. carinicauda BMP-7, AYC12382.1; E. sinensis BMP-7, AWV66904.1; S. paramamosain BMP-7, AOZ56940.1; H. sapiens BMP-7, NM_001719.3; G. gallus BMP-7, XM_417496.6; O. mossambicus BMP-7, FN565169.2; X. laevis BMP-7, NP_001079934.1.
FcBMP-7蛋白的系统进化树分析结果见图5。中国明对虾与凡纳滨对虾首先聚为一支,随后与脊尾白虾、中华绒螯蟹聚为一个甲壳类分支,然后与软体动物如文蛤、三角帆蚌等分支共同形成一个大的无脊椎动物分支,整个无脊椎动物分支又与脊椎动物分支共同组成完整的进化树,可见FcBMP-7蛋白在进化树中与中国明对虾的进化地位和分类阶元基本一致。
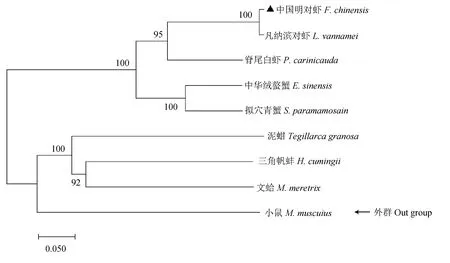
图5 基于邻接法构建的FcBMP-7 蛋白与其他已知物种BMP-7蛋白的系统进化树Fig.5 Phylogenetic tree of FcBMP-7 protein and other known species BMP-7 protein based on the neighbor-joining method
2.3 FcBMP-7基因的表达模式分析
2.3.1 FcBMP-7基因在中国明对虾幼体不同发育时期的转录水平分析
实时荧光定量PCR分析了FcBMP-7基因在中国明对虾幼体发育不同时期的表达情况,结果见图6。FcBMP-7基因在中国明对虾各幼体发育阶段均有表达,但表达水平不同,其中在原肠期表达最低,从无节幼体阶段开始表达量显著增加,糠虾幼体阶段达到最高,仔虾阶段表达量显著降低。
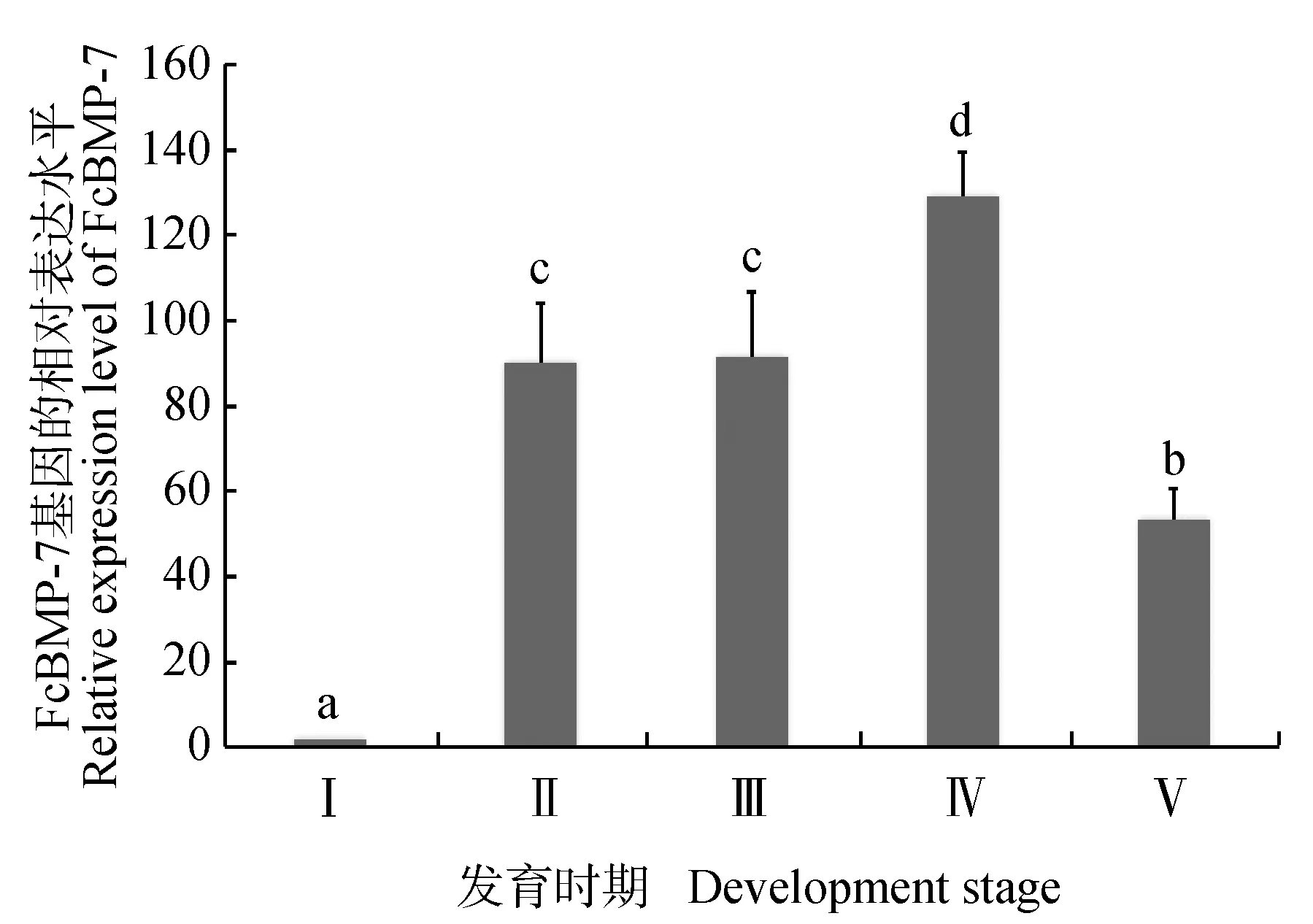
图6 FcBMP-7基因在中国明对虾幼体发育不同时期的转录表达水平Fig.6 The relative expression level of FcBMP-7 gene in Chinese shrimp F. chinensis at different development stagesⅠ.原肠期;Ⅱ.无节幼体期;Ⅲ.溞状幼体期;Ⅳ.糠虾幼体期;Ⅴ.仔虾期;不同字母表示组间存在显著性差异(P<0.05).Ⅰ.gastrula stage; Ⅱ.nauplius stage; Ⅲ.zoea stage; Ⅳ.mysis stage; Ⅴ.post-larval stage; means with different letters are significant differences among groups (P<0.05).
2.3.2 FcBMP-7基因在中国明对虾2种规格幼虾不同组织中的转录水平
FcBMP-7基因在中国明对虾2种规格幼虾不同组织中的表达结果见图7。FcBMP-7基因的表达量在2种幼虾大部分组织部位间存在显著差异,除了肝胰腺和鳃组织外,在 (1.55±0.12) g幼虾中FcBMP-7基因的表达量显著高于(3.95±0.08) g幼虾。在(3.95±0.08) g幼虾不同组织中,表达量最高的是鳃,其次是胃、游泳足和步足,表达量最少的是肌肉;在(1.55±0.12) g幼虾不同组织中,表达量最高的是鳃和游泳足,其次是步足和胃,表达量最少的是肝胰腺。

图7 FcBMP-7基因在2种规格虾不同组织中的表达水平Fig.7 The expression levels of FcBMP-7 gene in different tissues of large prawn and small juvenile shrimpA.肝胰腺;B.肌肉;C.肠道;D.心脏;E.胃;F.步足;G.游泳足;H.鳃; “*”表示两组之间存在显著性差异(P<0.05);“**”表示两组之间存在显著性差异(P< 0.01);“***”表示两组之间存在显著性差异(P<0.001).A.hepatopancreas; B.muscle; C.intestine; D.heart; E.stomach; F.periopod; G.pleopod; H.gill; “*” indicates significant difference between the two groups (P<0.05); “**” indicates significant difference between the two groups (P<0.01); “***” indicates significant difference between the two groups (P<0.001).
3.1 FcBMP-7基因的生物信息学分析
BMP-7蛋白作为TGF-β蛋白超家族中的一员,是Ozkaynak等[25]于牛成骨蛋白提取物中分离的一种新型转化生长因子。笔者通过cDNA末端快速扩增技术扩增克隆得到了FcBMP-7基因全长cDNA,对FcBMP-7蛋白质结构域的分析发现,该蛋白的信号肽长度为31位氨基酸,在羧基末端有一个重要的TGF-β多功能结构域(第307~413位氨基酸),即成熟肽,该结构域可控制多种细胞的增殖、生长、分化等功能。信号肽与成熟肽之间的第32~307位氨基酸称为前肽。与其他已知物种的BMP-7蛋白序列比对分析后发现,在此TGF-β保守结构域中,有7个高度保守的半胱氨酸残基,与人BMP-7蛋白单体的半胱氨酸残基结构相同, 可以形成3对链内二硫键, 剩余的1个半胱氨酸参与链间二硫键形成[26]。以上分析表明,FcBMP-7蛋白含有TGF-β蛋白超家族的特征序列,是TGF-β蛋白超家族中的多功能分泌性蛋白。对中国明对虾BMP-7蛋白与其他物种BMP-7蛋白的氨基酸序列进行聚类分析并构建系统进化树发现,即使经过漫长的进化和演变,对虾类BMP-7蛋白和脊椎动物BMP-7蛋白仍具有较高的同源性,表明该基因在整个动物界可能保持着相似的生理功能。
3.2 FcBMP-7基因的表达情况分析
近年来,对BMP-7基因的研究多集中在临床疾病[27-28]、软骨形成[29-31]以及各组织纤维化[32-33]等,对其在肌肉生长和发育中作用的研究相对较少[34]。为探明BMP-7基因在虾类肌肉生长发育中作用,笔者分析了FcBMP-7基因在中国明对虾早期发育不同阶段以及生长速率具有显著差异的2种规格幼虾不同组织中的表达水平。
3.2.1 FcBMP-7基因在中国明对虾幼体不同发育时期的转录水平
本试验中国明对虾幼体不同发育时期的表达结果显示,FcBMP-7基因在中国明对虾早期发育的5个时期均有表达,特别是在无节幼体时期开始显著高表达,在糠虾幼体期显著增加并达到表达高峰,仔虾期表达量逐渐降低。中国明对虾作为甲壳类的典型动物,具有其独特的发育模式,主要是肌肉系统的生成和完善。中国明对虾的肌肉从无节幼体时期开始陆续生成[35-36],糠虾幼体期是肌肉生成的重要阶段,直到仔虾期形成了较为完善的肌肉系统[37]。肌肉生成的过程与FcBMP-7基因的表达变化规律一致,由此推测FcBMP-7基因可能在对虾早期变态期的肌肉发生过程中发挥一定的作用,有待于后期进行深入的研究进行验证。
3.2.2 FcBMP-7基因在中国明对虾2种规格幼虾不同组织中的转录水平
通过分析中国明对虾2种规格幼虾不同组织的表达结果发现,FcBMP-7基因在中国明对虾的各组织中广泛性表达,表明FcBMP-7基因并不具备组织特异性表达特征,可能在对虾体内具有多种生理功能。值得注意的是,(3.95±0.08) g幼虾FcBMP-7基因的表达量除了在肝胰腺中高于(1.55±0.12) g幼虾外,在其他组织部位中的表达量均低于(1.55±0.12) g组幼虾(鳃差异不显著),与BMP-7基因在拟穴青蟹卵黄发育中的表达量随着卵黄的成熟而呈逐渐降低的趋势[38]相符合,并且存在明显的组织差异性。Yan等[39]对比分析了与本试验相同的2种规格幼虾不同组织部位(肌肉、心脏、步足、游泳足、鳃)肌纤维的数量与直径,结果显示,在肌肉、游泳足和步足等组织部位中,两组间肌纤维的数量与直径差异显著。虽然(3.95±0.08) g幼虾的肌纤维数量明显少于(1.55±0.12) g幼虾,但是前者肌肉纤维的直径明显大于后者。综合分析上述结果显示:FcBMP-7基因可能参与调控肌纤维的数量与生长,作为调节因子参与了中国明对虾幼虾的肌肉生长过程。
本试验结果初步表明,FcBMP-7基因在中国明对虾肌肉的发生及生长过程中具有一定的表达水平,推测FcBMP-7基因可能作为调节因子参与了中国明对虾幼虾肌肉发生和幼虾肌肉生长过程。该结果可为进一步研究FcBMP-7基因的功能提供重要的理论资料,同时为解析中国明对虾肌肉生长发育的调控机制提供线索。
猜你喜欢幼虾幼体对虾对虾养殖弱势群体的管理当代水产(2022年8期)2022-09-20黄海近岸潮汐锋海域蟹类幼体日间垂直迁移特征❋中国海洋大学学报(自然科学版)(2022年9期)2022-09-05对虾吃料慢的原因分析和处理当代水产(2022年5期)2022-06-05不同环境因素对克氏原螯虾后代幼虾生长状况的影响江苏农业科学(2022年4期)2022-03-11对虾免疫增强剂研究进展当代水产(2021年8期)2021-11-04对虾常见环境性疾病的防治当代水产(2021年4期)2021-07-20饵料类型、遮蔽比例与养殖密度对红螯螯虾幼虾生长的影响科学养鱼(2021年3期)2021-04-16探究“胚是新植物的幼体”实验创新生物学通报(2021年5期)2021-03-16奇异海蟑螂Ligia exotica幼体的生长特征研究海洋通报(2020年3期)2020-10-20不同隐蔽物环境下红螯螯虾幼虾的生长研究科学养鱼(2020年5期)2020-06-18